引言
Kcat 是转数 - 即每个酶位点每单位时间将多少底物分子转化为产物的数值。在分析底物浓度与反应速率曲线时,若已知酶位点的浓度,可使用 Kcat 代替 Vmax 进行拟合。
模型
Y = Et*kcat*X/(Km + X)
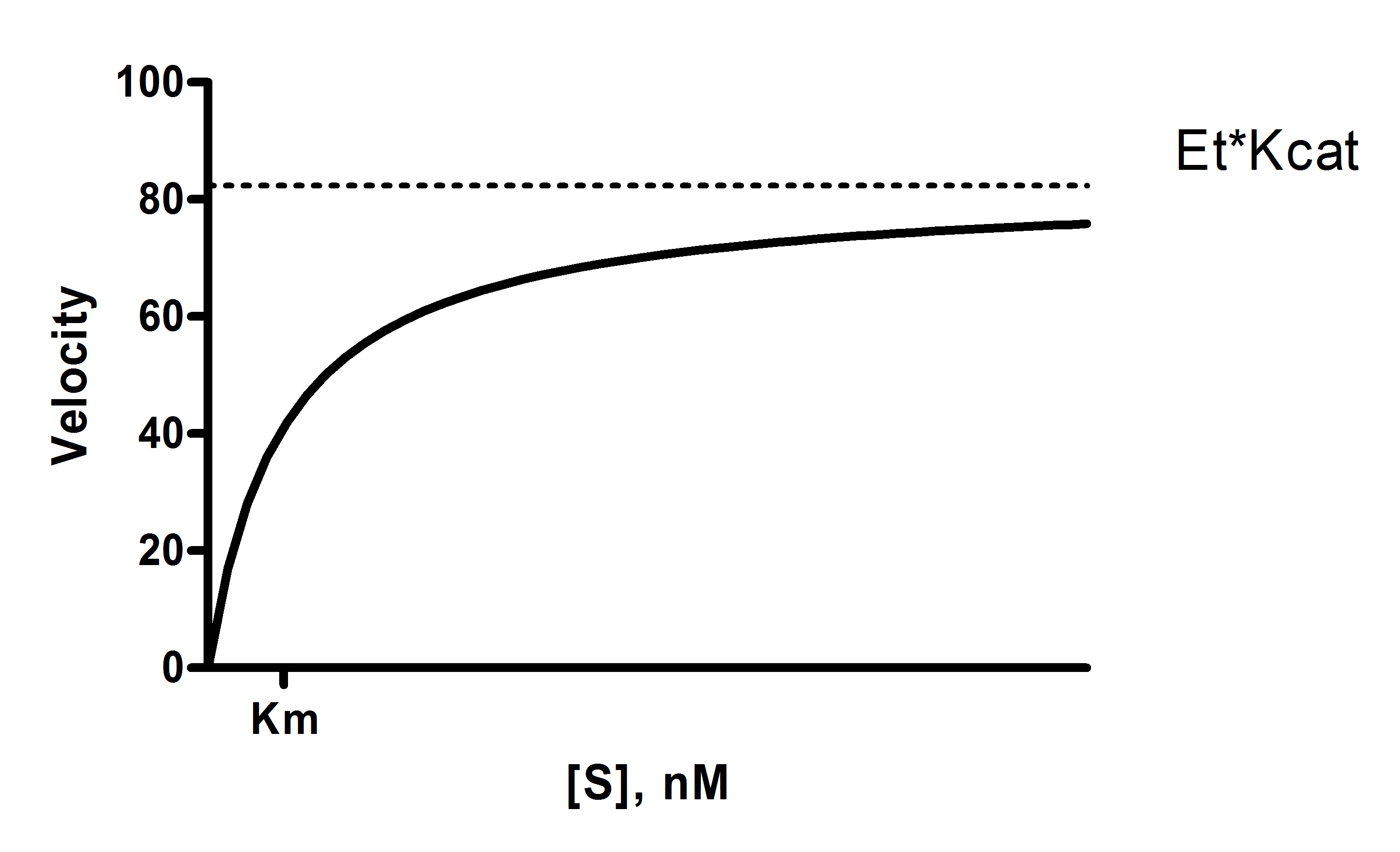
X 是底物浓度。
Y 为酶速率。
kcat 是转数,即每个酶位点每单位时间将底物转化为产物的次数。该值以 Y 轴时间单位的倒数表示。例如,若 Y 单位为每分钟微摩尔底物,则 kcat 即为每个催化位点每分钟产生的底物分子数。
Km 是米氏常数,单位与 X 相同。它是达到酶速率半最大值所需的底物浓度。
Et 是酶催化位点的浓度。如果酶由多个亚基组成,请注意 Et 表示催化位点的浓度,该值可能大于酶分子的浓度。您输入的 Y 值是以浓度/时间单位表示的酶速率。Et 必须使用相同的浓度单位输入(而时间单位由 kcat 定义)。
Vmax 是最大酶速率,单位与 Y 相同。它在上方的模型中并未直接显示。它是酶在极高底物浓度下外推得到的速率,因此几乎总是高于实验中测得的任何速率。其计算方法是 Et 乘以 kcat。
与米氏-门捷列夫模型的关系
上图所示曲线与米氏-门登模型定义的曲线完全一致。无论使用哪个模型拟合数据,所得曲线均完全相同,且Km值也完全一致。
米氏-门登模型求得的是 Vmax,即外推至极高底物浓度时的最大酶速率。其单位与您输入 Y 值(酶活性)时所用的单位相同。 通常,将其表示为(或转换为)摩尔/分钟/毫克蛋白质非常直观。Vmax 由存在的酶位点数量(Et)以及酶将底物转化为产物的速率(kcat)决定。
若已知加入实验中的酶位点浓度(Et),则可利用上述模型拟合催化常数 Kcat。
计算 Kcat 时,浓度单位会相互抵消,因此 Kcat 以倒数时间单位表示。它即转数 - 即每个酶位点每单位时间将底物分子转化为产物的数量。
使用 Prism 拟合模型
1.创建一个XY数据表。将底物浓度输入X列,酶速率输入Y列。如果有多个实验条件,将第一个条件置于A列,第二个置于B列,依此类推。您也可以选择Prism的示例数据:酶动力学 - 米氏-门登方程。
2.输入数据后,点击“分析”,选择“非线性回归”,选择“酶动力学方程”面板,并选择Kcat。
3.您必须根据其他实验结果将Et约束为一个常数值。要约束Et的值,请转到非线性回归对话框的“约束”选项卡,确保Et旁边的下拉菜单设置为“等于常数”,并输入该值。对于示例数据,请将Et的值输入为100。
若未知 Et 的数值,则无法拟合 Kcat,而应拟合 Vmax。Prism 无法同时拟合 Kcat 和 Et,因为这两个参数相互关联,且底物-速率曲线无法提供它们各自的具体数值。
4.根据示例数据,Prism 报告 Km= 5.886,其 95% 置信区间为 3.933 至 7.839。kcat 的控制数据为 13.53,其 95% 置信区间为 11.97 至 15.09。
注释
•参见所有酶动力学分析的假设列表。
•该方程拟合的曲线与拟合 Vmax 的方程完全一致,而非与转数 Kcat 相关。
•